微生物农药球孢白僵菌的发酵动力学研究毕业论文
2020-06-14 16:14:46
摘 要
本课题将对球孢白僵菌进行发酵,建立动力学模型,从表征上探讨其代谢水平差异。实验结果表明:用Monod模型、Logistic模型及Gompertz模型对白僵菌的发酵过程进行研究,其适用度分别为R2:0.8169、0.9982及0.9891,所以适用性最好的为Logistic模型; 对白僵菌发酵过程的菌体生长、底物消耗及产物生成过程进行分析,建立了白僵菌菌体生长的Logistic模型、产物生成的Luedeking-Piret和底物消耗的Luededing-Piret-like模型;在3 L发酵罐中对建立的三种动力学模型进行适用性验证,结果证明三种模型都较好的适用于白僵菌发酵的中试放大过程。这些动力学数据均可为球孢白僵菌优化、培养、工业化生产提供数据基础。
关键词:球孢白僵菌 发酵 动力学
Study on Fermentation Kinetics of Beauveria bassiana with Microorganism
Abstract
In this paper, we will ferment Beeleum bassiana to establish a dynamic model. Fr om the characterization, we explore its metabolic level differences. The results showed that the fermentation process of Beauveria bassiana was studied by Monod model, Logistic model and Gompertz model. The applicability was R2: 0.8169, 0.9982 and 0.9891, so the best applicability was Logistic model. The results showed that the logistic model of Beauveria bassiana was established, the Luedeking-Piret and the Luededing-Piret-like model were used for the production of the product. In the 3L fermentor, the results show that all three models are suitable for the mid - scale process of Beauveria bassiana fermentation. These kinetic data can provide a data basis for the optimization, cultivation and industrial production of Beauveria bassiana.
Keywords: Beauveria bassiana; fermentation; dynamics
目录
摘要 Ⅰ
ABSTRACT Ⅱ
第一章 文献综述 3
1.1 前言 3
1.2 球孢白僵菌的研究进展 4
1.3 球孢白僵菌的应用 5
1.4 本课题的研究目的与意义 6
第二章 球孢白僵菌发酵动力学研究 7
2.1 实验材料及方法 7
2.1.1 实验菌种 7
2.1.2 培养基 7
2.1.3 仪器设备 8
2.2 实验方法 8
2.2.1 菌株培养 8
2.2.2 分析检测方法 9
第三章 结果与讨论 10
3.1 葡萄糖标准曲线 11
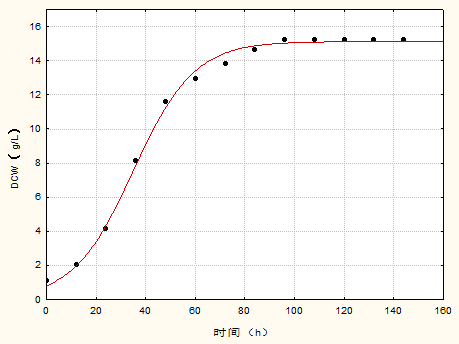
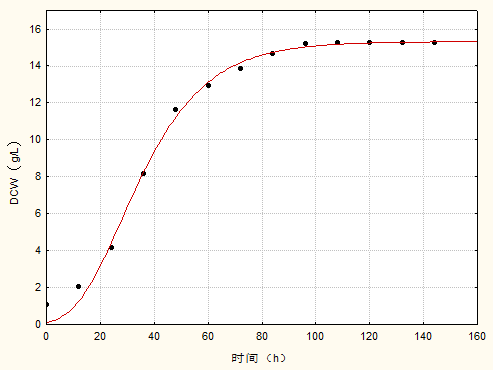
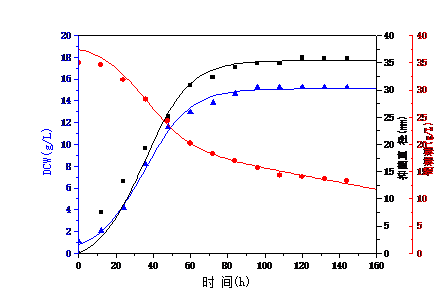
3.2 菌体生长动力学模型 11
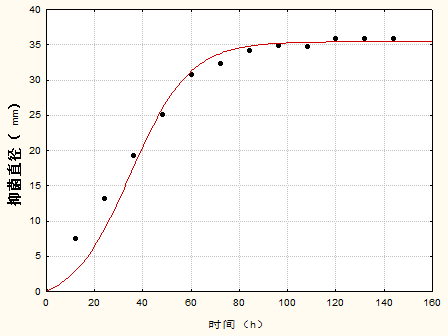
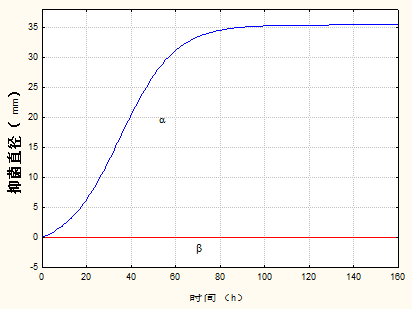
3.3 抑菌产物合成动力学模型 14
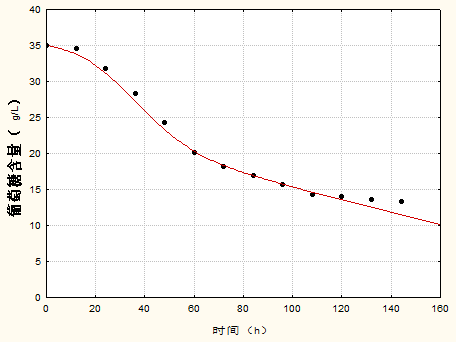
3.4 底物消耗动力学模型 16
3.5 动力学模型验证 17
第四章 结论与展望 20
4.1 结论 20
4.2 展望 20
参考文献 20
致 谢 20
第一章 文献综述
1.1 前言
球孢白僵菌是一种重要的广谱性昆虫病原真菌,在真菌分类学上属于半知菌亚门,丝孢纲,丛梗孢目,丛梗孢科,白僵菌属。球孢白僵菌菌落呈绒状、丛卷毛至粉状,有时呈绳索状,但很少形成孢梗索[1]。菌落培养初期多呈白色,稍后变成淡黄色,偶成淡红色;背面无色,或淡黄色至粉红色。分生孢子梗多着生在营养菌丝上,粗1-2μm,产孢细胞常浓密簇生(有时轮生或单生)于气生菌丝、稍有分化的分生孢子梗或膨大的泡囊(柄细胞)上,球形至瓶形,颈部明显延长成粗1μm、长达20μm 的产孢轴,轴上有小齿突,形成膝状 (“之”字形弯曲)的小齿状产孢轴。产孢细胞和泡囊常增生,在分生孢子梗或菌丝上聚成球状至卵状的相当密实的孢子头。分生孢子单孢,透明,薄壁,无厚垣孢子。
球孢白僵菌可通过分生孢子、芽生孢子、节孢子、内生孢子和菌丝体的断裂等多种方式进行无性增殖,其生活史类型属多孢型。在孢子萌发和菌丝生长过程中,通过芽管与芽管、芽管与菌丝或菌丝间的融合,不同基因型的细胞核可能会存在于同一细胞或菌丝体内形成复合型的重组子,即异核体。王成树[2]等依据球孢白僵菌28S rRNA 基因D11 域中内含子差异,扩增了117 个球孢白僵菌菌株,发现有17%的野生菌株以异核体的形式存在,其形成及频率分布与环境生态条件有一定相关性。白僵菌在虫体及培养基上皆以无性繁殖完成世代交替。关于白僵菌有性型的研究很多,但长期无定论,假散囊菌和头囊菌、虫草属都曾被疑为白僵菌的有性型。
球孢白僵菌是一种寄生真菌,昆虫是它最好的培养基。白僵菌的寄生范围极广,能侵染15 个目149个科的700多种昆虫及6个科的10余种蜱螨类。根据野外调查越冬昆虫发现,因白僵菌致死的昆虫占真菌致病总数的21%。白僵菌对于寄主的侵染方式可以分为两类,一种为体表侵入,另一种为直接由寄主食入而进入寄主身体,然后在寄主体内萌发生长直至把虫体中的营养消耗殆尽,其侵染以体表侵入虫体为主。感染的途径因昆虫种类、虫态、环境条件不同而有差别。白僵菌对寄主的侵染同其他虫生真菌一样,一般要经历10 个阶段,即:1.分生孢子附着于寄主体表;2.孢子在体表萌发;3.芽管穿透表皮进入寄主体腔;4.菌丝在血腔内生长;5.产生有毒代谢物;6.寄主死亡;7.菌丝侵入寄主的所有器官;8.菌丝穿出体表;9.在寄主体表重新形成孢子;10.分生孢子扩散。其中只要经历前4-5 个阶段,就可以使昆虫致病。金凯等[3]曾以草丁膦抗性基因bar 为筛选标记,构建了组成性表达绿色荧光蛋白基因egfp 的载体pBG,并成功转入球孢白僵菌,通过荧光观察清楚地检测到病原菌在昆虫体表的附着、菌丝穿透体壁以及菌丝从寄主体内长出等侵染过程。
1.2 球孢白僵菌的研究进展
随着大量的全基因组序列的测序完成,真菌生物学将进入功能基因组时代。目前,大约有14 种,真菌的基因组测序完成,而且还有至少100 种真菌基因组测序计划在进行之中。涉及球孢白僵菌基因工程研究的报道目前主要体现在三个方面:一是将球孢白僵菌作为宿主细胞接受外源基因的试验,并进而研究外源基因的稳定性;二是将球孢白僵菌的毒力因子(如Prl 基因)导入杀虫真菌受体菌,构建永久型表达的菌株,以提高真菌的杀虫效率;三是使用带有外源基因的稳定转化子进行释放试验,以验证工程菌株的环境稳定性及与当地菌株的基因交流情况。昆虫真菌分子生物学和生物技术研究起步稍晚,但近年来某些关键技术已取得突破。自从1995年Bundoek 等[4]首次成功地对酿酒酵母菌进行根癌农杆菌介导的遗传转化后,该方法已经在子囊菌、担子菌、接合菌中的几十种真菌上获得了成功。我国在国际上首次建立了球孢白僵菌(B.bassiana)的农杆菌转化体系,并从该菌中克隆了类枯草杆菌蛋白酶基因(Bbpr1)、几丁质酶基因(Bbchit1)等。如张永军等[5]将构建了超量表达类枯草杆菌蛋白酶基因CDEP-1的球孢白僵菌菌株Bb0062-15-4-CDEP-1,发现其发酵产物比野生菌株具有明显的杀桃蚜增效作用。
1.3 球孢白僵菌的应用
上世纪六十年代以后,人们逐渐认识到化学杀虫剂对生态环境的破坏性以及残留污染对人类自身健康的影响,便开始努力发展有效的替代防治方法,以利植物保护的可持续发展。球孢白僵菌作为一类重要的天敌生物资源,因其易于生产、寄主范围广、侵染期长、不伤害人和其他脊椎动物,尤其是其可贵的流行潜力而倍受人们的关注。长期以来,许多生物防治工作者对于真菌杀虫剂进行深入而又广泛的研究,并获得了巨大的成功。自1997年以来,欧美国家的公司先后登记注册了球孢白僵菌的系列制剂[6],如Mycotrol-ES、Myeotrol-WP、BotaniGard-ES、CornGard-ES、 CornGard-G(美国)、Ago Biocontrol Bassiana 50(哥伦比亚)、Ostrinil(法国)、Beauveria Schweizer(瑞典)、Engerlingspliz (德国) 等产品主要用于防治各种不允许存在化学农药。残留的高附加值的有机蔬菜瓜果等经济作物上的蚜虫、粉虱等刺吸式口器害虫、地下害虫、草原蝗虫及玉米螟、蔗螟等。
从20 世纪50 年代开始,球孢白僵菌已被应用于防治玉米螟、大豆食心虫和甘薯叶甲,20 世纪70 年代林业上应用球孢白僵菌大面积防治松毛虫并获得了明显成功。目前生产上应用球孢白僵菌防治松毛虫、玉米螟和水稻叶蝉等,每年防治面积达50 万hm2 左右,是世界上最成功的生物防治实例[7]。现在中国已成为世界上生产和使用白僵菌最多的国家。尤其是利用球孢自僵菌防治松毛虫的经验与技术令全世界瞩目。
相关图片展示:
课题毕业论文、开题报告、任务书、外文翻译、程序设计、图纸设计等资料可联系客服协助查找。


